خوش آمدید!
با عضویت در انجمن تک رمان از مزایای(چاپ کتاب،منتشر کردن رمان و...به صورت رایگان، خدمات ویراستاری، نقد و...)بهرمند شوید. با ما بهترینها را تجربه کنید.☆
همین حالا عضویتت رو قطعی کن!You are using an out of date browser. It may not display this or other websites correctly.
You should upgrade or use an alternative browser.
You should upgrade or use an alternative browser.
زیست شناسی ویروس ها
- نویسنده موضوع atena.vd
- تاریخ شروع
- پاسخها 34
- بازدیدها 1K
- Tagged users هیچ
ساعت تک رمان
مقاله ای از ویکی پدیا
___________________________
ویروس

ویروس موزائیک تنباکو:RNA احاطه شده توسط کپسید
ویروس قطعهای از نوکلئیک اسید است که درون یک پوشش پروتئینی (کپسید) محصور شدهاست. ویروسها از باکتریها بسیار کوچکتر هستند و اغلب با میکروسکوپ الکترونی قابل مشاهدهاند. ویروس با استفاده از امکانات سلولی میزبان تکثیر میشود و فعالیت اصلی یاختههای میزبان را مختل میکند. ویروسها بسیاری از جانوران و گیاهان و باکتریها را مبتلا میکنند اما فقط برخی از آنها انسانها را بیمار میکنند. همچنین ویروسها تنها در محیط خنثی در سلولهای زنده تکثیر میشوند و انگل اجباری داخل سلولی میباشند. چون ویروسها همهٔ ویژگیهای حیات را به غیر از فاز ژنتیک ندارند، زیست شناسان آنها را زنده نمیدانند. ویروسها رشد نمیکنند، حالت هومئوستازی ندارند و به دلیل نداشتن آنزیمهای اصلی واکنشهای متابولیسمی در آنها رخ نمیدهد. فقط دو آنزیم در آنها یافت میشود، آنزیم ATPآز و آنزیم رو نوشت بردار معکوس (مخصوص RNA دارها).
دانشمندان از مدتها پیش اینطور استدلال کردهاند که ویروسها غیر زنده هستند و فقط از ترکیب بیتهای DNA و RNA با سلولهای دیگر تشکیل شدهاند. در واقع بر اساس هر چیز دیگری که ما آنها را میشناسیم میتوانیم خصوصیات موجود زنده را شرح دهیم که یک ویروس نمیتواند این خصوصیات را داشته باشد. بسیاری از فرایندهای حیات مثل توانایی سوخت و ساز وجود دارند که ویروسها انجام نمیدهند. به نظر میرسد که ویروسها فقط قادر به انجام یکی از فرایندهای حیات هستند که آن تولید مثل است؛ ویروسهای فردی را حتی نمیتوان به عنوان یک ساختار دانست، یعنی حتی برای خواندن DNA و RNA آنها و ساخت یک ویروس جدید، نیاز به پروتینها میباشد. آنها به یک سلول حمله میکنند و ابزارهای ژنتیکی آن را برای خودشان میربایند.
اما در دهه گذشته تحولات ویروسشناسی شروع به فاش کردن اطلاعاتی کردهاست که نشان میدهد ویروسها ممکن است موجود زنده باشند. یکی از این اکتشافات میمی ویروسها (mimiviruses) بودند؛ ویروسهای غول پیکری با کتابخانههای ژنومی بزرگی که حتی از برخی از باکتریها نیز بزرگتر بودند. برخی از ویروسها مانند ویروس ابولا دارای تعداد کمی از ژن هفتم است. برخی از این غولها دارای ژنهای پروتئینهای هستند که برای ساخت یک ویروس جدید لازم و ضروری هستند. بر اساس این موضوع، ناشناخته ماندن این قضیه دلیلی بر قرار دادن آن ویروسها در طبقهبندی موجودات غیر زنده است
ویروسها ممکن است کروی، مارپیچی یا چند وجهی باشند. یک ویژگی مخصوص ویروسهای جانوری این است که یک پوششی از ج*ن*س غشا ی سلولی (پروتئین، لیپید و گلیکو پروتئین) دارند. این پوشش را از میزبان قبلی خود بدست میآورند و این بدان جهت است که ویروسهای جانوری از طریق آندوسیتوز وارد میزبان میشوند. ویروسهایی که به باکتریها حمله میکنند، باکتریوفاژ نام دارند که از سایر ویروسهای گیاهی و جانوری پیچیده ترند. این بدان علت است که شامل دو پوشش چند وجهی (به عنوان کپسید) و پوشش مارپیچی (به عنوان دم) میباشند.
محتویات
___________________________
ویروس
ویروس موزائیک تنباکو:RNA احاطه شده توسط کپسید
ویروس قطعهای از نوکلئیک اسید است که درون یک پوشش پروتئینی (کپسید) محصور شدهاست. ویروسها از باکتریها بسیار کوچکتر هستند و اغلب با میکروسکوپ الکترونی قابل مشاهدهاند. ویروس با استفاده از امکانات سلولی میزبان تکثیر میشود و فعالیت اصلی یاختههای میزبان را مختل میکند. ویروسها بسیاری از جانوران و گیاهان و باکتریها را مبتلا میکنند اما فقط برخی از آنها انسانها را بیمار میکنند. همچنین ویروسها تنها در محیط خنثی در سلولهای زنده تکثیر میشوند و انگل اجباری داخل سلولی میباشند. چون ویروسها همهٔ ویژگیهای حیات را به غیر از فاز ژنتیک ندارند، زیست شناسان آنها را زنده نمیدانند. ویروسها رشد نمیکنند، حالت هومئوستازی ندارند و به دلیل نداشتن آنزیمهای اصلی واکنشهای متابولیسمی در آنها رخ نمیدهد. فقط دو آنزیم در آنها یافت میشود، آنزیم ATPآز و آنزیم رو نوشت بردار معکوس (مخصوص RNA دارها).
دانشمندان از مدتها پیش اینطور استدلال کردهاند که ویروسها غیر زنده هستند و فقط از ترکیب بیتهای DNA و RNA با سلولهای دیگر تشکیل شدهاند. در واقع بر اساس هر چیز دیگری که ما آنها را میشناسیم میتوانیم خصوصیات موجود زنده را شرح دهیم که یک ویروس نمیتواند این خصوصیات را داشته باشد. بسیاری از فرایندهای حیات مثل توانایی سوخت و ساز وجود دارند که ویروسها انجام نمیدهند. به نظر میرسد که ویروسها فقط قادر به انجام یکی از فرایندهای حیات هستند که آن تولید مثل است؛ ویروسهای فردی را حتی نمیتوان به عنوان یک ساختار دانست، یعنی حتی برای خواندن DNA و RNA آنها و ساخت یک ویروس جدید، نیاز به پروتینها میباشد. آنها به یک سلول حمله میکنند و ابزارهای ژنتیکی آن را برای خودشان میربایند.
اما در دهه گذشته تحولات ویروسشناسی شروع به فاش کردن اطلاعاتی کردهاست که نشان میدهد ویروسها ممکن است موجود زنده باشند. یکی از این اکتشافات میمی ویروسها (mimiviruses) بودند؛ ویروسهای غول پیکری با کتابخانههای ژنومی بزرگی که حتی از برخی از باکتریها نیز بزرگتر بودند. برخی از ویروسها مانند ویروس ابولا دارای تعداد کمی از ژن هفتم است. برخی از این غولها دارای ژنهای پروتئینهای هستند که برای ساخت یک ویروس جدید لازم و ضروری هستند. بر اساس این موضوع، ناشناخته ماندن این قضیه دلیلی بر قرار دادن آن ویروسها در طبقهبندی موجودات غیر زنده است
ویروسها ممکن است کروی، مارپیچی یا چند وجهی باشند. یک ویژگی مخصوص ویروسهای جانوری این است که یک پوششی از ج*ن*س غشا ی سلولی (پروتئین، لیپید و گلیکو پروتئین) دارند. این پوشش را از میزبان قبلی خود بدست میآورند و این بدان جهت است که ویروسهای جانوری از طریق آندوسیتوز وارد میزبان میشوند. ویروسهایی که به باکتریها حمله میکنند، باکتریوفاژ نام دارند که از سایر ویروسهای گیاهی و جانوری پیچیده ترند. این بدان علت است که شامل دو پوشش چند وجهی (به عنوان کپسید) و پوشش مارپیچی (به عنوان دم) میباشند.
محتویات
- ۱ کشف ویروسها
- ۲ ساختار ویروسها
- ۳ تکثیر ویروسها
- ۴ تکثیر ویروسها مراحل مختلفی را طی میکند که به شرح آن میپردازیم
- ۴.۱ جذب و اتصال
- ۴.۲ نفوذ یا Penetration (ورود به سلول)
- ۴.۳ پوشش برداری
- ۴.۴ ناپدید شدن
- ۴.۵ تکثیر یا بیوسنتز
- ۴.۶ تکمیل شدن (کامل شدن)
- ۴.۷ خروج ویروس از سلول
- ۵ ژنتیک ویروسها
- ۶ تغییرات دو جانبه ژنتیکی ویروسها
- ۶.۱ نوترکیبی
- ۶.۲ فعال شدن متقاطع
- ۶.۳ دوباره فعال شدن افزایشی
- ۶.۴ تغییرات دو جانبه غیر ژنتیکی
- ۶.۵ اختلاط ژنوتیپی
- ۶.۶ تداخل (انترفرانس)
- ۷ جستارهای وابسته
- ۸ منابع
کشف ویروسها
در پایان قرن ۱۹ میلادی دانشمندان به دنبال یافتن عامل بیماری موزائیک تنباکو بودند. دیمیتری ایوانفسکی درسال۱۸۵۸ توانست قابل انتقال بودن عامل بیماری موزانیک توتون را ثابت کند. او پس از عبور دادن عصاره برگهای گیاه عفونت زده از صافی غیرقابل عبور برای باکتریها، آن را بروی برگهای گیاه سالم توتون پخش کرده و بیماری موزانیک توتون را انتقال داده ایجاد بیماری درگیاه سالم نمود. آنان دریافتند که از گیاه آلوده مولدی قادر است که گیاهان سالم را بیمار کند؛ بنابراین عامل بیماری هرچه که بود از باکتریها بسیار کوچکتر بود. در سال ۱۹۳۵ زیستشناسی به نام وندل استنلی توانست ویروس موزاییک تنباکو را تخلیص کند. ویروس خالص شده اگر چه به شکل بلور بود اما باز هم میتوانست گیاهان سالم را بیمار کند. چون تبلور یک از ویژگیهای مواد شیمیایی است بنابراین میتوان نتیجه گرفت که ویروس داخل یاختهها موجودی زنده و خارج یاختهها یک ماده شیمایی است نه یک موجود زنده. همچنین بایجرینیگ پدر علم ویروسشناسی است.
انجمن رمان نویسی
در پایان قرن ۱۹ میلادی دانشمندان به دنبال یافتن عامل بیماری موزائیک تنباکو بودند. دیمیتری ایوانفسکی درسال۱۸۵۸ توانست قابل انتقال بودن عامل بیماری موزانیک توتون را ثابت کند. او پس از عبور دادن عصاره برگهای گیاه عفونت زده از صافی غیرقابل عبور برای باکتریها، آن را بروی برگهای گیاه سالم توتون پخش کرده و بیماری موزانیک توتون را انتقال داده ایجاد بیماری درگیاه سالم نمود. آنان دریافتند که از گیاه آلوده مولدی قادر است که گیاهان سالم را بیمار کند؛ بنابراین عامل بیماری هرچه که بود از باکتریها بسیار کوچکتر بود. در سال ۱۹۳۵ زیستشناسی به نام وندل استنلی توانست ویروس موزاییک تنباکو را تخلیص کند. ویروس خالص شده اگر چه به شکل بلور بود اما باز هم میتوانست گیاهان سالم را بیمار کند. چون تبلور یک از ویژگیهای مواد شیمیایی است بنابراین میتوان نتیجه گرفت که ویروس داخل یاختهها موجودی زنده و خارج یاختهها یک ماده شیمایی است نه یک موجود زنده. همچنین بایجرینیگ پدر علم ویروسشناسی است.
انجمن رمان نویسی
ساختار ویروسها

طرحی ساده از ساختار یک ویروس
تفاوت این ویروسها با کلیه میکروبهای شناخته شده، آنان را در زیستشناسی میکروبی از مقام و جایگاه ویژه و ردهبندی جداگانهای برخوردار کردهاست. ساختمان ویروسها محدود است به ژنوم و پوششی ار پروتئین به دور آن. در شماری از ویروسها لایهای از لیپو پروتئین و لیپوپلی ساکارید نیز مضافاً برای حفاظت پوشش پروتئینی به دور آن قرار میگیرد. (کپسید و پوشش ویروس را ببینید) ویروسها برای تکثیر باید به داخل سلولی نفوذ کرده برنامه تولیدی سلول را بنفع خود تغییرداده و کارگاه پروتئینسازی سلول را همهجانبه برای تولید پروتئینها و آنزیمهای لازم بکار گیرند. ویروسها در عین کوچکی با داشتن ژنهای کافی مراحل بسیار پیچیده تکثیر خود را درسلول میزبان یا به گروگان گرفته شده به سهولت هدایت و کنترل میکنند و انگل وار تکثیر میشوند. ژنوم ویروسها عبارت است از نوعی اسید نوکلئیک که بر مبنای نوع آن، ویروسها به دو دسته تقسیم میشوند. ویروسهای حاوی "آر ان اِى" ر یبونوکلئیک اسید RNA و ویروسهای حاوی "دیانای" دئوکسی ریبونوکلئیک اسید. DNA عفونت زایی ویروسها فقط و فقط حاصل فعالیت داخل سلولی اسید نوکلئیک یا ژنوم آنهاست به این جهت درخارج سلول ویروس و ژنومش عاری از هرگونه فعالیت بوده و یک مولکول بزرگ محسوب میشود. ژنوم بسیاری از ویروسها به صورت رشته یا کلاف داخل پوشش پروتئینشان که اختصاصاًکاپسید کپسیدنامیده میشود قراردارد. این پوشش پروتیینی یکپارچه نبوده بلکه از ذرات یا واحدهای کوچکتری به نام کاپسومر ساخته میشود. کاپسومرها پس از بهم پیوستن کاپسیدرا تشکیل میدهند و فرم مخصوص آن بستگی به نوع ویروس و ساختار آن دارد. در برخی ویروسها ژنوم به صورت مارپیچی سنتز شده و کاپسومرها آن را بطور منظم پوشش میدهند، طوریکه نوکلئوکاپسید ویروس حالت فنری یا هلیکال بخود میگیرد. علاوه بر این ژنوم و کاپسید (نوکلئوکاپسید) بیش از نیمی از ویروسها دارای غلاف (به انگلیسی: Envelope) از لیپوپروتئین و لیپوپولی ساکارید ند. در این مورد هم ویروسها به دو دسته تقسیم میشوند. گروه غلافدار و گروه بدون غلاف. غلاف ویروسها از تنوع بسیاری برخورداراست که بهطور مفصل به آن خواهیم پرداخت. اندازه، شکل، حجم و ساختمان ویروسها بسیار متنوع و متفاوت است. ویروسها برخی کروی، بعضی مخروطی و بسیاری از اشکال هندسی مانند برخوردارند، ازآن جمله میتوان فرم مکعبی یا پلی هدرون را مثال زد. قطر ویروس ها میتواند از عدد ۲۰ نانومتر (کوچکترین) تا ۴۵۰ نانومتر (بزرگترین) متغییر باشد. ویروسهای بزرگ مانند ویروس آبله از ساختمانی نسبتاً پیچیده یا مرکب (complex) برخوردارند و همچون باکتریها ابداً از صافی (فیلترچینی) عبور نمیکنند. درصورتیکه مابقی توانایی گذر کردن از این صافی را دارند. ویروسها فقط به علت داشتن اطلاعات ژنتیکی برای تولیدمثل به جانداران شبیهاند. در واقع ویروسها انگل اجباری هستند. دستاوردهای فوق تائیدیست برای اینکه، برخلاف کلیه موجودات زنده اعم از تک سلولی یا بسیار سلولی وپروکاریونت و یوکاریونتها، ویروسها فاقد ساختمان سلولی بوده عاری از هرگونه فعل و انفعالات شیمیایی، آنزیمی و متابولیسم سلولی اند وبا فقدان ملزومات مشابه سازی، هرگز قادر به تکثیر و مشابهسازی خود نیستند. آنها را باید خارج از سلول زنده یک ماکرومولکول مرکب یا یک واحد عفونت زا بهشمارآورد. آنها با توجه به اینکه ماکرومولکولهای مرکبند یعنی دارای زنجیره اسید نوکلئیک و حفاظ پروتئینی و احتمالاً غلافی از لیپوپروتئین و لیپولی ساکارید اند، آمادهاند به محض تماس با سلول میزبان و پس از مراحل جذب، دخول و آزادی ژنوم دستگاه پروتئینسازی میزبان را در اختیار گرفته و مراحل کلی تکثیرشان را به کمک سیستمهای سلولی (ترانسکریپسیون و ترانسلاسیون) ترتیب دهند. سلولها اعم از از پروکاریونت و یوکاریونتها میتوانند سلول میزبان اجباری ویروسها گردند. هرنوع سلولی پذیرای هر ویروسی نبوده، انحصارا ویروس آشنا بخود را پذیرا میباشد.
انجمن رمان نویسی
طرحی ساده از ساختار یک ویروس
تفاوت این ویروسها با کلیه میکروبهای شناخته شده، آنان را در زیستشناسی میکروبی از مقام و جایگاه ویژه و ردهبندی جداگانهای برخوردار کردهاست. ساختمان ویروسها محدود است به ژنوم و پوششی ار پروتئین به دور آن. در شماری از ویروسها لایهای از لیپو پروتئین و لیپوپلی ساکارید نیز مضافاً برای حفاظت پوشش پروتئینی به دور آن قرار میگیرد. (کپسید و پوشش ویروس را ببینید) ویروسها برای تکثیر باید به داخل سلولی نفوذ کرده برنامه تولیدی سلول را بنفع خود تغییرداده و کارگاه پروتئینسازی سلول را همهجانبه برای تولید پروتئینها و آنزیمهای لازم بکار گیرند. ویروسها در عین کوچکی با داشتن ژنهای کافی مراحل بسیار پیچیده تکثیر خود را درسلول میزبان یا به گروگان گرفته شده به سهولت هدایت و کنترل میکنند و انگل وار تکثیر میشوند. ژنوم ویروسها عبارت است از نوعی اسید نوکلئیک که بر مبنای نوع آن، ویروسها به دو دسته تقسیم میشوند. ویروسهای حاوی "آر ان اِى" ر یبونوکلئیک اسید RNA و ویروسهای حاوی "دیانای" دئوکسی ریبونوکلئیک اسید. DNA عفونت زایی ویروسها فقط و فقط حاصل فعالیت داخل سلولی اسید نوکلئیک یا ژنوم آنهاست به این جهت درخارج سلول ویروس و ژنومش عاری از هرگونه فعالیت بوده و یک مولکول بزرگ محسوب میشود. ژنوم بسیاری از ویروسها به صورت رشته یا کلاف داخل پوشش پروتئینشان که اختصاصاًکاپسید کپسیدنامیده میشود قراردارد. این پوشش پروتیینی یکپارچه نبوده بلکه از ذرات یا واحدهای کوچکتری به نام کاپسومر ساخته میشود. کاپسومرها پس از بهم پیوستن کاپسیدرا تشکیل میدهند و فرم مخصوص آن بستگی به نوع ویروس و ساختار آن دارد. در برخی ویروسها ژنوم به صورت مارپیچی سنتز شده و کاپسومرها آن را بطور منظم پوشش میدهند، طوریکه نوکلئوکاپسید ویروس حالت فنری یا هلیکال بخود میگیرد. علاوه بر این ژنوم و کاپسید (نوکلئوکاپسید) بیش از نیمی از ویروسها دارای غلاف (به انگلیسی: Envelope) از لیپوپروتئین و لیپوپولی ساکارید ند. در این مورد هم ویروسها به دو دسته تقسیم میشوند. گروه غلافدار و گروه بدون غلاف. غلاف ویروسها از تنوع بسیاری برخورداراست که بهطور مفصل به آن خواهیم پرداخت. اندازه، شکل، حجم و ساختمان ویروسها بسیار متنوع و متفاوت است. ویروسها برخی کروی، بعضی مخروطی و بسیاری از اشکال هندسی مانند برخوردارند، ازآن جمله میتوان فرم مکعبی یا پلی هدرون را مثال زد. قطر ویروس ها میتواند از عدد ۲۰ نانومتر (کوچکترین) تا ۴۵۰ نانومتر (بزرگترین) متغییر باشد. ویروسهای بزرگ مانند ویروس آبله از ساختمانی نسبتاً پیچیده یا مرکب (complex) برخوردارند و همچون باکتریها ابداً از صافی (فیلترچینی) عبور نمیکنند. درصورتیکه مابقی توانایی گذر کردن از این صافی را دارند. ویروسها فقط به علت داشتن اطلاعات ژنتیکی برای تولیدمثل به جانداران شبیهاند. در واقع ویروسها انگل اجباری هستند. دستاوردهای فوق تائیدیست برای اینکه، برخلاف کلیه موجودات زنده اعم از تک سلولی یا بسیار سلولی وپروکاریونت و یوکاریونتها، ویروسها فاقد ساختمان سلولی بوده عاری از هرگونه فعل و انفعالات شیمیایی، آنزیمی و متابولیسم سلولی اند وبا فقدان ملزومات مشابه سازی، هرگز قادر به تکثیر و مشابهسازی خود نیستند. آنها را باید خارج از سلول زنده یک ماکرومولکول مرکب یا یک واحد عفونت زا بهشمارآورد. آنها با توجه به اینکه ماکرومولکولهای مرکبند یعنی دارای زنجیره اسید نوکلئیک و حفاظ پروتئینی و احتمالاً غلافی از لیپوپروتئین و لیپولی ساکارید اند، آمادهاند به محض تماس با سلول میزبان و پس از مراحل جذب، دخول و آزادی ژنوم دستگاه پروتئینسازی میزبان را در اختیار گرفته و مراحل کلی تکثیرشان را به کمک سیستمهای سلولی (ترانسکریپسیون و ترانسلاسیون) ترتیب دهند. سلولها اعم از از پروکاریونت و یوکاریونتها میتوانند سلول میزبان اجباری ویروسها گردند. هرنوع سلولی پذیرای هر ویروسی نبوده، انحصارا ویروس آشنا بخود را پذیرا میباشد.
انجمن رمان نویسی
تکثیر ویروسها
چرخه همانندسازی ویروسها از آلوده شدن سلول میزبان آغاز میشود و با رهاسازی ذرات ویروسی ایجادشده بالغ خاتمه مییابد.
ویروسها به علتِ فقدانِ ساختمانِ سلولی و هرگونه متابولیسم و فعل و انفعالِ شیمیایی، قادر به مشابهسازی خود نبوده و برای این عمل میباید به داخل سلولی حساس راه یافته و محتاجِ انرژی و دستگاهِ پروتئین سازیِ سلولِ زنده میباشند. انتقالِ ویروس به داخلِ سلول فقط توسطِ سلول امکانپذیر است و این عمل فقط توسطِ سلولِ حساس و حاملِ گیرندههای (رسپتورهای) آشنا به ویروس انجام میگیرد. سلولهایی که این نوع گیرندهها را برای جذبِ ویروس آماده دارند احتمالاً میتوانند به انتقالِ انواعِ مختلفِ ویروسها بپردازند، در غیر اینصورت سلول در مقابلِ ویروس مقاوم بوده و هرگونه تماسی با ویروس بینتیجه خواهد بود. با دخول به سلول و پوشش برداری توسطِ آنزیمهایِ سلول، فعالیتِ اسیدنوکلئیک ویروس آغاز میگردد. اسیدنوکلئیکِ ویروس ژنهای کافی برایِ مهارنِمودنِ متابولیسمِ سلول میزبان را دارا بوده و به کمکِ آن احتیاجاتِ فعل و انفعالاتِ شیمی حیاتی، برای تکثیرِ خود را توسطِ سلولِ میزبان تأمین میکند. پس از آلوده شدنِ سلولِ میزبان تکثیر ویروسها میتواند در دو نوع چرخه انجام گیرد:
چرخه لیزوژنی:
گاهی ویروس پس از دخول به سلول پس از طی مراحل اولیه و آزادشدنِ ژنوم یا اسیدنوکلئیک، به عوضِ تولید ژنوم و پروتئین ویروس، خود را درونِ کروموزومِ میزبان جای میدهد که در این حالت به ان پرو ویروس میگویند با هر بار تقسیم سلولی، پرو ویروس نیز تقسیم میشود. در این نوع چرخه بدونِ آنکه سلولِ میزبان تخریب شود ژنومِ ویروسی تکثیر پیدا میکند که گاهی این مرحله بدون آنکه سلولِ میزبان تخریب شود به تولیدِ ویروسِ کامل پرداخته و ویروسهای نوزاد را به خارجِ سلول هدایت میکند.
چرخه لیتیک:
دراین مرحله سلولِ میزبان پس از انجامِ تکثیر ویروس به کلی تخریب شده از این راه ویروسهای زیادی از سلول میزبان آزاد میشوند.
انجمن رمان نویسی
چرخه همانندسازی ویروسها از آلوده شدن سلول میزبان آغاز میشود و با رهاسازی ذرات ویروسی ایجادشده بالغ خاتمه مییابد.
ویروسها به علتِ فقدانِ ساختمانِ سلولی و هرگونه متابولیسم و فعل و انفعالِ شیمیایی، قادر به مشابهسازی خود نبوده و برای این عمل میباید به داخل سلولی حساس راه یافته و محتاجِ انرژی و دستگاهِ پروتئین سازیِ سلولِ زنده میباشند. انتقالِ ویروس به داخلِ سلول فقط توسطِ سلول امکانپذیر است و این عمل فقط توسطِ سلولِ حساس و حاملِ گیرندههای (رسپتورهای) آشنا به ویروس انجام میگیرد. سلولهایی که این نوع گیرندهها را برای جذبِ ویروس آماده دارند احتمالاً میتوانند به انتقالِ انواعِ مختلفِ ویروسها بپردازند، در غیر اینصورت سلول در مقابلِ ویروس مقاوم بوده و هرگونه تماسی با ویروس بینتیجه خواهد بود. با دخول به سلول و پوشش برداری توسطِ آنزیمهایِ سلول، فعالیتِ اسیدنوکلئیک ویروس آغاز میگردد. اسیدنوکلئیکِ ویروس ژنهای کافی برایِ مهارنِمودنِ متابولیسمِ سلول میزبان را دارا بوده و به کمکِ آن احتیاجاتِ فعل و انفعالاتِ شیمی حیاتی، برای تکثیرِ خود را توسطِ سلولِ میزبان تأمین میکند. پس از آلوده شدنِ سلولِ میزبان تکثیر ویروسها میتواند در دو نوع چرخه انجام گیرد:
چرخه لیزوژنی:
گاهی ویروس پس از دخول به سلول پس از طی مراحل اولیه و آزادشدنِ ژنوم یا اسیدنوکلئیک، به عوضِ تولید ژنوم و پروتئین ویروس، خود را درونِ کروموزومِ میزبان جای میدهد که در این حالت به ان پرو ویروس میگویند با هر بار تقسیم سلولی، پرو ویروس نیز تقسیم میشود. در این نوع چرخه بدونِ آنکه سلولِ میزبان تخریب شود ژنومِ ویروسی تکثیر پیدا میکند که گاهی این مرحله بدون آنکه سلولِ میزبان تخریب شود به تولیدِ ویروسِ کامل پرداخته و ویروسهای نوزاد را به خارجِ سلول هدایت میکند.
چرخه لیتیک:
دراین مرحله سلولِ میزبان پس از انجامِ تکثیر ویروس به کلی تخریب شده از این راه ویروسهای زیادی از سلول میزبان آزاد میشوند.
انجمن رمان نویسی
تکثیر ویروسها مراحل مختلفی را طی میکند که به شرح آن میپردازیم
جذب و اتصال
ویروس با گیرندههای در غشاء سلولی تماس پیدا نموده (Reseptor) و سبب جذب و اتصال ویروس به سلول شده و شروع عفونت پایهگذاری میشود.این اتصال ویروس به رسپتور و وجود رسپتورهای مذکور در بافتهای خاص باعث اختصاصیت ویروس به بافتهای خاص و ایجاد بیماریهای مشخص در همان سلولهای بافتی شدهاست . حرارت محیط، پی اچ مناسب، نیروی الکترواستاتیک و وجود نمکهای معدنی درجذب ویروس و اتصال به رسپتورهای سطح سلول از اهمیت زیادی برخوردارند.اتصال ویروس به رسپتور اتصالی ضعیف و غیر کووالانسی است . نوع رسپتورها برای ویروسهای مختلف یکسان نیست برای مثال ویروس فلج اطفال جذب PVRو ویروس آنفلوآنزا جذب سیالیک اسید سلولهای تنفسی میگردد. همچنین رسپتور ویروس HIV مولکول CD4 و رسپتور ویروس ابشتن بار مولکولCD21 و رسپتور رینوویروس مولکول ICAM-1 میباشد . هر کدام از گیرندهها عملکردی طبیعی در خدمت به ب*دن دارند و ویروس از آنها سو استفاده میکند ،به طوری که CD4 در سلولهای لنفوسیت T نقشی به عنوان کمک محرک برای شناسایی و پاسخ به آنتی ژنهای بیگانه متصل به MHC برعهده دارد .
نفوذ یا Penetration (ورود به سلول)
بعد از اتصال ویروس به سطح سلول با تغییراتی در رسپتور سلولی ویروس به دنبال راهی برای ورود به سلول میباشد .در کل ورود ویرو یا از طریف فیوژن و یا از طریق اندوسیتوزصورت میگیرد . ویروسهای دارای پوشش اغلب از طریق فیوژن و با کمک پروتئینهای ویروسی که نقش در فیوژن دارند وارد سلول میشوند . سلولهای بدون پوشینه نیز از طریق اندوسیتوز وارد سلول میشوند ،که هرکدام انواعی از اندوسیتوز شامل اندوسیتوز وابسته به کلاترین، اندوسیتوز وابسته با کائولین ، اندوسیتوز مستقل از کلاترین و کائولین و مارپینوسیتوز میباشد .
پوشش برداری
پس از دخول ویروس به سلول، در داخل سیتوپلاسم و تحت تأثیر آنزیمهای داخل سلولی پوشش پروتئینی ویروس از هم گسسته و اسید نوکلئیک آزاد میگردد. از آغاز تا این مرحله را میتوان با کمک میکروسکوپ الکترونی تغییرات ویروس را دنبال کرده، مشاهده نمود.
ناپدید شدن
اسید نوکلئیک آزاد شده ویروس به قطعات کوچکتری تقسیم شده و پس از اندک زمانی کمترین اثری از ژنوم ویروس قابل رؤیت نخواهدبود. از این پس اسید نوکلئیک ویروس فرمانروای سلول عفونت زده بوده و کنترل دستگاه پروتئینسازی سلول را برای تکثیر خویش هدایت میکند. در این مراحل میتوان ویروس را یک واحد زنده تلقی نمود یا به عبارت دیگر اسید نوکلئیک ویروس یک واحد زنده محسوب میشود.
تکثیر یا بیوسنتز
با سنتز و ساخت و ساز آنزیمهای لازمه تکثیر، فعالیت اسید نوکلئیک ویروس در داخل سیتوپلاسم سلولی شروع شده، آنزیمها فعالیت خود را برای جلوگیری از تولید پروتئین و اسید نوکلئیک سلولی آغاز میکنند. در بعضی از موارد تولید پروتئین و اسید نوکلئیک سلولی بهطور کلی متوقف نشده و فقط به نحو کنترل شده محدود میگردد. به هر حال مراحل ترانسلاسیون و ترانسکریپسیون برای بیوسنتز و رپلیکاسیون ویروس با تولید و ساخت پروتئین و اسید نوکلئیک ویروس در مراکز مشخصی آغاز میگردد. مراکز تولید با توجه به نوع ویروس هر کدام در گوشهای از سلول، گاهی درداخل سیتوپلاسم و گاهی در داخل هسته سلول تشکیل شده، آماده تهیه و تولید ویروس و مرحله بعدی آن میگردند.
تکمیل شدن (کامل شدن)
پس از تولید و سنتز پروتئین و اسید نوکلئیک ویروس در مراکز مشخص. اسید نوکلئیک ویروس در داخل پروتئین محافظ جایگرفته و کار تکمیل شدن ویروس روند مختص به خود را پیش میگیرد. تکمیل شدن ویروسها بهطور عمومی یکسان نبوده و بهطور خصوصی انجام میگیرد. تکمیل شدن ویروسها ی غلافدار یا بدون غلاف و آنان که دارای ساختمان پیچیده (کمپلکس) هستند یا دارای نوکلئوکاپسید فنری (هلیکال) اند کاملاً متفاوت از هم بوده و هر کدام سیر تکاملی دیگری طی کردهاند. بطور خلاصه میتوان گفت که ویروسهای کروی یا بعبارت دیگر با کاپسید ایکوزاهدرون با بهم پیوستن کاپسومرها و جای گرفتن اسید نوکلئیک در داخل آن کامل میشوند.. ویروسهایی که نوکلئوکاپسید فنری دارند قبلاً به روی اسید نوکلئیک پوششی از کاپسومرها قرار گرفته و فرم فنری خود را هنگامی بدست میآورند که ویروس برای خروج از سلول آماده میشود. ویروسهایی که دارای ساختمان پیچیده کمپلکس هستند تکمیل شدنشان بسادگی دیگر ویروسها نبوده و بهطوریکه بررسیها نشان میدهند اسید نوکلئیک و پروتئین حافظ و فیلامانهای سطح خارجی آنها و لیپیدی که در ساختمانشان بکار میرود بطور کلی در محل دیگری سنتز شده و ویروس در همان محل فرم میگیرد و کامل میشود غلاف ویروسهای غلافدار، آنهایی که در داخل هسته سلول تکثیر میشوند پس از خروج از هسته و آنهایی که در سیتوپلاسم سلول تکمیل میشود هنگام خروج غلافی از غشاء سیتوپلاسمی بدورشان پیچیده شده تکمیل میگردند.
خروج ویروس از سلول
در مورد خروج ویروسها از سلول دو روش مشاهده میگردد. در بعضی موارد ویروس پس از اینکه تکمیل شد بسته به نوع آن و نوع سلول، با متلاشی شدن سلول میزبان، ویروس آزاد میشود. در اینگونه موارد، پس از انهدام سلول مقدار زیادی ویروس ناگهان آزاد میشود. در بعضی موارد د یگر ویروس بدون اینکه سلول منهدم شود به خارج از سلول رها میگردد. در این موارد گاهی ویروس همانند جوانهای از سلول جدا میشود. در مورد ویروسهای غلافدار همزمان غلافشان به دورشان پیچیده شده و از سلول بخارج راه میابند.
انجمن رمان نویسی
جذب و اتصال
ویروس با گیرندههای در غشاء سلولی تماس پیدا نموده (Reseptor) و سبب جذب و اتصال ویروس به سلول شده و شروع عفونت پایهگذاری میشود.این اتصال ویروس به رسپتور و وجود رسپتورهای مذکور در بافتهای خاص باعث اختصاصیت ویروس به بافتهای خاص و ایجاد بیماریهای مشخص در همان سلولهای بافتی شدهاست . حرارت محیط، پی اچ مناسب، نیروی الکترواستاتیک و وجود نمکهای معدنی درجذب ویروس و اتصال به رسپتورهای سطح سلول از اهمیت زیادی برخوردارند.اتصال ویروس به رسپتور اتصالی ضعیف و غیر کووالانسی است . نوع رسپتورها برای ویروسهای مختلف یکسان نیست برای مثال ویروس فلج اطفال جذب PVRو ویروس آنفلوآنزا جذب سیالیک اسید سلولهای تنفسی میگردد. همچنین رسپتور ویروس HIV مولکول CD4 و رسپتور ویروس ابشتن بار مولکولCD21 و رسپتور رینوویروس مولکول ICAM-1 میباشد . هر کدام از گیرندهها عملکردی طبیعی در خدمت به ب*دن دارند و ویروس از آنها سو استفاده میکند ،به طوری که CD4 در سلولهای لنفوسیت T نقشی به عنوان کمک محرک برای شناسایی و پاسخ به آنتی ژنهای بیگانه متصل به MHC برعهده دارد .
نفوذ یا Penetration (ورود به سلول)
بعد از اتصال ویروس به سطح سلول با تغییراتی در رسپتور سلولی ویروس به دنبال راهی برای ورود به سلول میباشد .در کل ورود ویرو یا از طریف فیوژن و یا از طریق اندوسیتوزصورت میگیرد . ویروسهای دارای پوشش اغلب از طریق فیوژن و با کمک پروتئینهای ویروسی که نقش در فیوژن دارند وارد سلول میشوند . سلولهای بدون پوشینه نیز از طریق اندوسیتوز وارد سلول میشوند ،که هرکدام انواعی از اندوسیتوز شامل اندوسیتوز وابسته به کلاترین، اندوسیتوز وابسته با کائولین ، اندوسیتوز مستقل از کلاترین و کائولین و مارپینوسیتوز میباشد .
پوشش برداری
پس از دخول ویروس به سلول، در داخل سیتوپلاسم و تحت تأثیر آنزیمهای داخل سلولی پوشش پروتئینی ویروس از هم گسسته و اسید نوکلئیک آزاد میگردد. از آغاز تا این مرحله را میتوان با کمک میکروسکوپ الکترونی تغییرات ویروس را دنبال کرده، مشاهده نمود.
ناپدید شدن
اسید نوکلئیک آزاد شده ویروس به قطعات کوچکتری تقسیم شده و پس از اندک زمانی کمترین اثری از ژنوم ویروس قابل رؤیت نخواهدبود. از این پس اسید نوکلئیک ویروس فرمانروای سلول عفونت زده بوده و کنترل دستگاه پروتئینسازی سلول را برای تکثیر خویش هدایت میکند. در این مراحل میتوان ویروس را یک واحد زنده تلقی نمود یا به عبارت دیگر اسید نوکلئیک ویروس یک واحد زنده محسوب میشود.
تکثیر یا بیوسنتز
با سنتز و ساخت و ساز آنزیمهای لازمه تکثیر، فعالیت اسید نوکلئیک ویروس در داخل سیتوپلاسم سلولی شروع شده، آنزیمها فعالیت خود را برای جلوگیری از تولید پروتئین و اسید نوکلئیک سلولی آغاز میکنند. در بعضی از موارد تولید پروتئین و اسید نوکلئیک سلولی بهطور کلی متوقف نشده و فقط به نحو کنترل شده محدود میگردد. به هر حال مراحل ترانسلاسیون و ترانسکریپسیون برای بیوسنتز و رپلیکاسیون ویروس با تولید و ساخت پروتئین و اسید نوکلئیک ویروس در مراکز مشخصی آغاز میگردد. مراکز تولید با توجه به نوع ویروس هر کدام در گوشهای از سلول، گاهی درداخل سیتوپلاسم و گاهی در داخل هسته سلول تشکیل شده، آماده تهیه و تولید ویروس و مرحله بعدی آن میگردند.
تکمیل شدن (کامل شدن)
پس از تولید و سنتز پروتئین و اسید نوکلئیک ویروس در مراکز مشخص. اسید نوکلئیک ویروس در داخل پروتئین محافظ جایگرفته و کار تکمیل شدن ویروس روند مختص به خود را پیش میگیرد. تکمیل شدن ویروسها بهطور عمومی یکسان نبوده و بهطور خصوصی انجام میگیرد. تکمیل شدن ویروسها ی غلافدار یا بدون غلاف و آنان که دارای ساختمان پیچیده (کمپلکس) هستند یا دارای نوکلئوکاپسید فنری (هلیکال) اند کاملاً متفاوت از هم بوده و هر کدام سیر تکاملی دیگری طی کردهاند. بطور خلاصه میتوان گفت که ویروسهای کروی یا بعبارت دیگر با کاپسید ایکوزاهدرون با بهم پیوستن کاپسومرها و جای گرفتن اسید نوکلئیک در داخل آن کامل میشوند.. ویروسهایی که نوکلئوکاپسید فنری دارند قبلاً به روی اسید نوکلئیک پوششی از کاپسومرها قرار گرفته و فرم فنری خود را هنگامی بدست میآورند که ویروس برای خروج از سلول آماده میشود. ویروسهایی که دارای ساختمان پیچیده کمپلکس هستند تکمیل شدنشان بسادگی دیگر ویروسها نبوده و بهطوریکه بررسیها نشان میدهند اسید نوکلئیک و پروتئین حافظ و فیلامانهای سطح خارجی آنها و لیپیدی که در ساختمانشان بکار میرود بطور کلی در محل دیگری سنتز شده و ویروس در همان محل فرم میگیرد و کامل میشود غلاف ویروسهای غلافدار، آنهایی که در داخل هسته سلول تکثیر میشوند پس از خروج از هسته و آنهایی که در سیتوپلاسم سلول تکمیل میشود هنگام خروج غلافی از غشاء سیتوپلاسمی بدورشان پیچیده شده تکمیل میگردند.
خروج ویروس از سلول
در مورد خروج ویروسها از سلول دو روش مشاهده میگردد. در بعضی موارد ویروس پس از اینکه تکمیل شد بسته به نوع آن و نوع سلول، با متلاشی شدن سلول میزبان، ویروس آزاد میشود. در اینگونه موارد، پس از انهدام سلول مقدار زیادی ویروس ناگهان آزاد میشود. در بعضی موارد د یگر ویروس بدون اینکه سلول منهدم شود به خارج از سلول رها میگردد. در این موارد گاهی ویروس همانند جوانهای از سلول جدا میشود. در مورد ویروسهای غلافدار همزمان غلافشان به دورشان پیچیده شده و از سلول بخارج راه میابند.
انجمن رمان نویسی
آخرین ویرایش:
ژنتیک ویروسها
ژنتیک ویروسهای انسانی و حیوانی تاکنون بطور کامل شناخته نشدهاست. علت اصلی این امر فقدان ساختمان سلولی و متابولیسم اختصاصی آنهاست. چون تکثیرشان کاملاً وابسته و گره خورده به سلول میزبان است، بررسی جزء به جزء آن دشوار و مراحل مختلف آن به سختی قابل پیگیری میباشد. از طرفی ویروسها از نظر ژنتیکی استوارتر از باکتریها بوده و تغییرات ژنتیکی در آنها به غیر از ویروسهای آنفلوآنزا و ایدز در سایر ویروسهای انسانی و حیوانی خیلی به ندرت اتفاق میافتد و به دست آوردن موتانتها یا رکمبینانتهای نوین بسیار مشکل است. از اینرو بررسی ژنتیک آنها بسادگی امکانپذیر نیست و دشواری آن در درمان بیماریهای ویروسی منعکس میگردد.
تغییرات دو جانبه ژنتیکی ویروسها
تغییرات دو جانبه ژنوم ویروسها هنگامی به وقوع میپیوندند که ژنوم دو ویروس فعال همزمان به داخل سلولی راه یافته و آن را عفونی نموده، سلول میزبان عمل تکثیر را در مورد هرد و با انجام رسانیده باشد. در اینگونه موارد احتمال تغییرات و تعویض دو جانبه ژنتیکی در میان دو ویروس بسیار زیاد است که در این موارد قسمتی از اسید نوکلئیک ویروس اول به اسید نوکلئیک ویروس دیگر پیوسته اجباراً و ژنهای جدیدی را همرا آورده به ژنهای اولیه اضافه میکند. در صورتیکه قسمتی از اسید نوکلئیک ویروس اول به اسید نوکلئیک ویروس دوم پیوسته و قسمتی از اسید نوکلئیک ویروس دوم به اسید نوکلئیک ویروس اول بپیوندد در این حالت ژنوم هر کدام قسمتی از دست میدهد و قسمتی دیگر به دست میآورد.
بطور کلی در اینگونه موارد میتوان احتمالات زیر را مشاهده نمود:
نوترکیبی
نوترکیبی، تعویض ترمیمی متقابل قسمتهایی از ژنوم دو ویروس فعال است که همزمان بداخل سلولی راه یافته و آن را عفونی نموده باشند. تعویض اتفاقی قسمتی از ژنوم دو ویروس فعال گاهی سبب افزایش ویرولانس یا کاهش آن و احتمالاً تغییراتی در آنتیژنهای ویروس به عمل میآید. زمانی حاصل آن بدست آوردن خصوصیات جدیدیست که ویروس تا بحال فاقد آن بوده وبا کسب آن به کلی تغییر ماهیت بدهد. رکمبیناسیون در ویروسهای آبله و آنفلوآنزا و فلج اطفال و باکتروفاژها گزارش شدهاست.
فعال شدن متقاطع
تلاقی ژنوم ویروس فعالی با ژنوم ویروس غیرفعالی که همزمان بداخل سلولی راه یافته و آن را عفونی نموده باشند گاهی منجر به تعویض متقاطع قسمتی از ژنوم آنها شده و هر دو ویروس حاصل این تغییرات خصوصیات جدیدی را کسب کرده و در نتیجه هر دو ویروس پدیدار و فعال میگردند. برای مثال جهت تهیه واکسن از ویروسیهایی که در سلولهای معینی قابل تکثیر نیستند میتوان آنها را به کمک این تغییرات ژنتیکی برای تکثیرشان در سلولهای مورد نظر فعال نمود.
دوباره فعال شدن افزایشی
زمانی است که دو ویروس غیرفعال به سلولی راه میابند و اسید نوکلئیک غیرفعال آن دو به تعمیر و ترمیم یکدیگر پرداخته در نتیجه یک ویروس فعال و دارای خصوصیات هر دو بوجود میآید. این امر و افزایش ژنتیکی متقابل که در این صورت اتفاق میافتد احتمال ویروسی فعال و خشن حاصل میگردد.
تغییرات دو جانبه غیر ژنتیکی
هنگامی دو ویروس همزمان به سلولی راه یابند و تکثیر شوند، اگر اطلاعات ژنتیکی ویروس اولی (ژنوتیپ) در کاپسید (فنوتیپ) ویروس دومی جایگزین شود اختلاط فنوتیپیک رخ میدهد. این امر موجب پیدایش دو ویروس جدید که بطور اتفاقی فنوتیپ و ژنوتیپ آنها تغییر یافته میگردد. این حالت پایدار نبوده، در صورتیکه هر کدام سلولی را عفونی سازند ویروسهای نوزاد فنوتیپ عاریه را از دست داده فنوتیپ اصلی خود را عیان خواهند نمود.
اختلاط ژنوتیپی
گاهی عفونت همزمان سلول با دو ویروس مختلف منجر به اختلاط ژنوتیپ آنها میگردد؛ بنابراین درصورتی که که یک ویروس از نظر ژنتیکی اطلاعات کامل دو ویروس مختلف را دربرداشته رد یعنی دو ژنوم مختلف در کاپسیدش جای گرفته باشند از اختلاط ژنوتیپ سخن میگوییم. در این گونه موارد نیز، ثبات ژنتیکی وجود ندارد. حاصل اولین تکثیر آن ویروس منجر به پدیدار شدن دو ویروس مختلف خواهند بود. این پدیده در ویروسهای پارامیکسو گزارش شدهاست
تداخل (انترفرانس)
تجربه نشان میدهد، گاهی کشت مای سلولی پس از عفونی شدن به وسیلهٔ ویروسی در قبال عفونت با ویروسهای دیگری نوعی مصونیت نشان میدهند. حتی اگر ویروس دیگری بداخل سلول نفوذ کند به تکثیر نمیشود. علت آن به احتمال زیاد ترشح انترفرون سلولی یا کنترل و هدایت متابولیسم سلول توسط ویروس عفونت زا و عدم امکان برای کنترل و هدایت سلول توسط ویروس دوم میباشد. عکس عمل انتر فرانس نیز امکانپذیر است. در این صورت سلولهای عفونی شده توسط ویروس یکم تکثیر ویروس دوم را تشدید و افزایش میدهند. احتمالاً علت این امر جلوگیری از ترشح انترفرون به کمک اطلاعات ژنتیکی ویروس دوم است. عکس انتر فرانس امکان دیگریست که با همکاری دو ویروس در جلوگیری از ادامه متابولیسم سلول نتیجه به نفع هر دو ویروس بوده مراحل تکثیر هر دو ویروس ممکن میسازد. گاهی هنگامیکه دو ویروس ناکامل و ناقص سلولی را عفونی میکنند، گرچه هیچکدام به تنهائی امکان را نداشته و احتمال تکثیرشان بسیار ناچیز است، اطلاعات ژنتیکی دو ویروس میتوانند مکمل یکدیگر شده وبا مهار و کنترل متابولیسم سلول هر دو آنها به تکثیر رسند.
انجمن رمان نویسی
ژنتیک ویروسهای انسانی و حیوانی تاکنون بطور کامل شناخته نشدهاست. علت اصلی این امر فقدان ساختمان سلولی و متابولیسم اختصاصی آنهاست. چون تکثیرشان کاملاً وابسته و گره خورده به سلول میزبان است، بررسی جزء به جزء آن دشوار و مراحل مختلف آن به سختی قابل پیگیری میباشد. از طرفی ویروسها از نظر ژنتیکی استوارتر از باکتریها بوده و تغییرات ژنتیکی در آنها به غیر از ویروسهای آنفلوآنزا و ایدز در سایر ویروسهای انسانی و حیوانی خیلی به ندرت اتفاق میافتد و به دست آوردن موتانتها یا رکمبینانتهای نوین بسیار مشکل است. از اینرو بررسی ژنتیک آنها بسادگی امکانپذیر نیست و دشواری آن در درمان بیماریهای ویروسی منعکس میگردد.
تغییرات دو جانبه ژنتیکی ویروسها
تغییرات دو جانبه ژنوم ویروسها هنگامی به وقوع میپیوندند که ژنوم دو ویروس فعال همزمان به داخل سلولی راه یافته و آن را عفونی نموده، سلول میزبان عمل تکثیر را در مورد هرد و با انجام رسانیده باشد. در اینگونه موارد احتمال تغییرات و تعویض دو جانبه ژنتیکی در میان دو ویروس بسیار زیاد است که در این موارد قسمتی از اسید نوکلئیک ویروس اول به اسید نوکلئیک ویروس دیگر پیوسته اجباراً و ژنهای جدیدی را همرا آورده به ژنهای اولیه اضافه میکند. در صورتیکه قسمتی از اسید نوکلئیک ویروس اول به اسید نوکلئیک ویروس دوم پیوسته و قسمتی از اسید نوکلئیک ویروس دوم به اسید نوکلئیک ویروس اول بپیوندد در این حالت ژنوم هر کدام قسمتی از دست میدهد و قسمتی دیگر به دست میآورد.
بطور کلی در اینگونه موارد میتوان احتمالات زیر را مشاهده نمود:
نوترکیبی
نوترکیبی، تعویض ترمیمی متقابل قسمتهایی از ژنوم دو ویروس فعال است که همزمان بداخل سلولی راه یافته و آن را عفونی نموده باشند. تعویض اتفاقی قسمتی از ژنوم دو ویروس فعال گاهی سبب افزایش ویرولانس یا کاهش آن و احتمالاً تغییراتی در آنتیژنهای ویروس به عمل میآید. زمانی حاصل آن بدست آوردن خصوصیات جدیدیست که ویروس تا بحال فاقد آن بوده وبا کسب آن به کلی تغییر ماهیت بدهد. رکمبیناسیون در ویروسهای آبله و آنفلوآنزا و فلج اطفال و باکتروفاژها گزارش شدهاست.
فعال شدن متقاطع
تلاقی ژنوم ویروس فعالی با ژنوم ویروس غیرفعالی که همزمان بداخل سلولی راه یافته و آن را عفونی نموده باشند گاهی منجر به تعویض متقاطع قسمتی از ژنوم آنها شده و هر دو ویروس حاصل این تغییرات خصوصیات جدیدی را کسب کرده و در نتیجه هر دو ویروس پدیدار و فعال میگردند. برای مثال جهت تهیه واکسن از ویروسیهایی که در سلولهای معینی قابل تکثیر نیستند میتوان آنها را به کمک این تغییرات ژنتیکی برای تکثیرشان در سلولهای مورد نظر فعال نمود.
دوباره فعال شدن افزایشی
زمانی است که دو ویروس غیرفعال به سلولی راه میابند و اسید نوکلئیک غیرفعال آن دو به تعمیر و ترمیم یکدیگر پرداخته در نتیجه یک ویروس فعال و دارای خصوصیات هر دو بوجود میآید. این امر و افزایش ژنتیکی متقابل که در این صورت اتفاق میافتد احتمال ویروسی فعال و خشن حاصل میگردد.
تغییرات دو جانبه غیر ژنتیکی
هنگامی دو ویروس همزمان به سلولی راه یابند و تکثیر شوند، اگر اطلاعات ژنتیکی ویروس اولی (ژنوتیپ) در کاپسید (فنوتیپ) ویروس دومی جایگزین شود اختلاط فنوتیپیک رخ میدهد. این امر موجب پیدایش دو ویروس جدید که بطور اتفاقی فنوتیپ و ژنوتیپ آنها تغییر یافته میگردد. این حالت پایدار نبوده، در صورتیکه هر کدام سلولی را عفونی سازند ویروسهای نوزاد فنوتیپ عاریه را از دست داده فنوتیپ اصلی خود را عیان خواهند نمود.
اختلاط ژنوتیپی
گاهی عفونت همزمان سلول با دو ویروس مختلف منجر به اختلاط ژنوتیپ آنها میگردد؛ بنابراین درصورتی که که یک ویروس از نظر ژنتیکی اطلاعات کامل دو ویروس مختلف را دربرداشته رد یعنی دو ژنوم مختلف در کاپسیدش جای گرفته باشند از اختلاط ژنوتیپ سخن میگوییم. در این گونه موارد نیز، ثبات ژنتیکی وجود ندارد. حاصل اولین تکثیر آن ویروس منجر به پدیدار شدن دو ویروس مختلف خواهند بود. این پدیده در ویروسهای پارامیکسو گزارش شدهاست
تداخل (انترفرانس)
تجربه نشان میدهد، گاهی کشت مای سلولی پس از عفونی شدن به وسیلهٔ ویروسی در قبال عفونت با ویروسهای دیگری نوعی مصونیت نشان میدهند. حتی اگر ویروس دیگری بداخل سلول نفوذ کند به تکثیر نمیشود. علت آن به احتمال زیاد ترشح انترفرون سلولی یا کنترل و هدایت متابولیسم سلول توسط ویروس عفونت زا و عدم امکان برای کنترل و هدایت سلول توسط ویروس دوم میباشد. عکس عمل انتر فرانس نیز امکانپذیر است. در این صورت سلولهای عفونی شده توسط ویروس یکم تکثیر ویروس دوم را تشدید و افزایش میدهند. احتمالاً علت این امر جلوگیری از ترشح انترفرون به کمک اطلاعات ژنتیکی ویروس دوم است. عکس انتر فرانس امکان دیگریست که با همکاری دو ویروس در جلوگیری از ادامه متابولیسم سلول نتیجه به نفع هر دو ویروس بوده مراحل تکثیر هر دو ویروس ممکن میسازد. گاهی هنگامیکه دو ویروس ناکامل و ناقص سلولی را عفونی میکنند، گرچه هیچکدام به تنهائی امکان را نداشته و احتمال تکثیرشان بسیار ناچیز است، اطلاعات ژنتیکی دو ویروس میتوانند مکمل یکدیگر شده وبا مهار و کنترل متابولیسم سلول هر دو آنها به تکثیر رسند.
انجمن رمان نویسی
آرانای ویروس
ویروس آرانای یا ویروس با آر.ان.ای مثبت نوعی از ویروسهاست که ژنوم یا ماده ژنتیکی آن از آرانای تشکیل شدهاست.[۱] مانند ویروس هپاتیت سی ، فلج اطفال ، سرخک ، سرماخوردگی و ابولا .
نوکلئیکاسید و ژنوم در ویروس آر.ان.ای معمولاً تکرشتهای و گاهی نیز جفترشتهای است و همچنین در دو شکل ممتد یا قطعهقطعه وجود دارد. مثلاً ویروس سرخک دارای آر.ان.ای ممتد و ویروس آنفلوآنزا دارای ژنوم آر.ان.ای قطعهقطعه است.
طبقهبندی
ژنوم در ویروس آر.ان.ای معمولاً تکرشتهای (ssRNA) و گاهی نیز جفترشتهای (dsRNA) است.
آر.ان.ای تکرشتهای براساس پولاریته تعریف شده و درصورت همقطب بودن پولاریته آر.ان.ای و پولاریته ام.آر.ان.ای (RNA پیامرسان) مثبت است. درحالت مقابل٬ اگر ایندو مکمل هم باشند پولاریته منفی محسوب میشود. ویروسهای پولاریتهمنفی برای رونویسی از آر.ان.ای پیامبر نیاز به آنزیم ویژهای بنام ترانسکریپتاز خواهند داشت.[۲]
بعضی از آرانای ویروسهای تکرشتهای که قادر به ایجاد عفونت در سلولهای جانوری هستند در داخل ذره ویروسی یک DNA پلیمراز وابسته به RNA به نام ترانس کریپتاز معکوس (Reverse transcriptase) دارند .
در هنگام ایجاد عفونت ژنوم RNA تک رشته ویروس با حدود ۱۰۰۰۰ نوکلئوتید همراه به آنزیم ترانس کریپتاز معکوس و یک tRNA به عنوان حامل آنزیم برداشته شده از سلول میزبان قبلی وارد سلول میزبان جدید میشود . tRNAتبدیل فوری RNA ویروسی به DNA دو رشتهای را تسهیل می نماید . آنزیم ترانس کریپتاز معکوس به عنوان DNA پلیمراز وابسته به RNA ابتدا سنتز یک رشته DNA مکمل RNA ویروسی را کاتالیز میکند . سپس با کمک محیط قلیایی NaOH رشته RNA در هیبرید DNA - RNA تخریب شده و سپس به کمک DNA پلی مراز I رشته دوم DNA ساخته میشود و تکثیر ژنوم ویروس آغاز میشود.
انجمن رمان نویسی
ویروس آرانای یا ویروس با آر.ان.ای مثبت نوعی از ویروسهاست که ژنوم یا ماده ژنتیکی آن از آرانای تشکیل شدهاست.[۱] مانند ویروس هپاتیت سی ، فلج اطفال ، سرخک ، سرماخوردگی و ابولا .
نوکلئیکاسید و ژنوم در ویروس آر.ان.ای معمولاً تکرشتهای و گاهی نیز جفترشتهای است و همچنین در دو شکل ممتد یا قطعهقطعه وجود دارد. مثلاً ویروس سرخک دارای آر.ان.ای ممتد و ویروس آنفلوآنزا دارای ژنوم آر.ان.ای قطعهقطعه است.
طبقهبندی
ژنوم در ویروس آر.ان.ای معمولاً تکرشتهای (ssRNA) و گاهی نیز جفترشتهای (dsRNA) است.
آر.ان.ای تکرشتهای براساس پولاریته تعریف شده و درصورت همقطب بودن پولاریته آر.ان.ای و پولاریته ام.آر.ان.ای (RNA پیامرسان) مثبت است. درحالت مقابل٬ اگر ایندو مکمل هم باشند پولاریته منفی محسوب میشود. ویروسهای پولاریتهمنفی برای رونویسی از آر.ان.ای پیامبر نیاز به آنزیم ویژهای بنام ترانسکریپتاز خواهند داشت.[۲]
بعضی از آرانای ویروسهای تکرشتهای که قادر به ایجاد عفونت در سلولهای جانوری هستند در داخل ذره ویروسی یک DNA پلیمراز وابسته به RNA به نام ترانس کریپتاز معکوس (Reverse transcriptase) دارند .
در هنگام ایجاد عفونت ژنوم RNA تک رشته ویروس با حدود ۱۰۰۰۰ نوکلئوتید همراه به آنزیم ترانس کریپتاز معکوس و یک tRNA به عنوان حامل آنزیم برداشته شده از سلول میزبان قبلی وارد سلول میزبان جدید میشود . tRNAتبدیل فوری RNA ویروسی به DNA دو رشتهای را تسهیل می نماید . آنزیم ترانس کریپتاز معکوس به عنوان DNA پلیمراز وابسته به RNA ابتدا سنتز یک رشته DNA مکمل RNA ویروسی را کاتالیز میکند . سپس با کمک محیط قلیایی NaOH رشته RNA در هیبرید DNA - RNA تخریب شده و سپس به کمک DNA پلی مراز I رشته دوم DNA ساخته میشود و تکثیر ژنوم ویروس آغاز میشود.
انجمن رمان نویسی
دیانای ویروس
دیانای ویروسها (DNA virus) گروهی از ویروسهایند که ژنوم یا ماده ژنتیکی آن از دیانای تشکیل شدهاست و تکثیر ماده ژنتیکی در این ویروسها توسط دیانای-پلیمراز وابسته به دیانای انجام میگیرد مانند ویروس تبخال ، آبله مرغان و آبله.
طبقه بندی
در طبقهبندی بالتیمور دیانای ویروسها به دو گروه تقسیم میشوند: گروه یک ویروس های با DNA دو رشته ای (dsDNA) و گروه دو دیانای تک رشته ای (ssDNA) . ویروسهای با دیانای دورشتهای که اکثریت هستند مانند آدنوویروس ، ویروس پاپیلوم انسانی ، سیتومگالوویروس و هرپس ویروسها . ویروسهای با دیانای تکرشتهای مانند پاروویروس.
دقت شود که در طبقهبندی بالتیمور هپادنوویروسها (مانند ویروس هپاتیت بی) هرچند دارای ژنوم دیانای دورشتهای حلقوی (cccDNA) هستند ولی چون همانندسازی ژنوم در این ویروسها از طریق آرانای پیامرسان (mRNA) انجام میگیرد (نه دیانای-پلیمراز ) لذا در گروه دیانای ویروسها قرار ندارند بلکه گروه هفتم (Group VII: Double-stranded DNA viruses that replicate through a single-stranded RNA intermediate) را تشکیل میدهند.
انجمن رمان نویسی
دیانای ویروسها (DNA virus) گروهی از ویروسهایند که ژنوم یا ماده ژنتیکی آن از دیانای تشکیل شدهاست و تکثیر ماده ژنتیکی در این ویروسها توسط دیانای-پلیمراز وابسته به دیانای انجام میگیرد مانند ویروس تبخال ، آبله مرغان و آبله.
طبقه بندی
در طبقهبندی بالتیمور دیانای ویروسها به دو گروه تقسیم میشوند: گروه یک ویروس های با DNA دو رشته ای (dsDNA) و گروه دو دیانای تک رشته ای (ssDNA) . ویروسهای با دیانای دورشتهای که اکثریت هستند مانند آدنوویروس ، ویروس پاپیلوم انسانی ، سیتومگالوویروس و هرپس ویروسها . ویروسهای با دیانای تکرشتهای مانند پاروویروس.
دقت شود که در طبقهبندی بالتیمور هپادنوویروسها (مانند ویروس هپاتیت بی) هرچند دارای ژنوم دیانای دورشتهای حلقوی (cccDNA) هستند ولی چون همانندسازی ژنوم در این ویروسها از طریق آرانای پیامرسان (mRNA) انجام میگیرد (نه دیانای-پلیمراز ) لذا در گروه دیانای ویروسها قرار ندارند بلکه گروه هفتم (Group VII: Double-stranded DNA viruses that replicate through a single-stranded RNA intermediate) را تشکیل میدهند.
انجمن رمان نویسی
بیماری ویروسی
نوشتار اصلی: ویروس
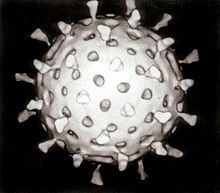
ویروسها موجودات بسیار کوچکی هستند که ساختار سلولی ندارند، به همین دلیل برای تولیدمثل باید وارد سلول زنده شوند و با استفاده از مواد موجود در سلول میزبان تولیدمثل کنند.
ویروس در بیرون از ب*دن هیچ گونه فعالیت زیستی ندارد.
محتویات
ویروسها تقریباً تمام موجودات زنده را مورد حمله قرار میدهند.
دفاع ب*دن در برابر ویروسها
هنگام حمله ویروسها به ب*دن، سلولهای مورد حمله مادهای پروتئینی به نام اینترفرون تولید میکنند. پس از پارهشدن سلولها و آزادشدن ویروسها، اینترفرون به سلولهای سالم میچسبد و آن سلولها را نسبت به ویروس بطور موقت مقاوم میکند.
موارد استفاده ویروسها
نوشتار اصلی: ویروس
ویروسها موجودات بسیار کوچکی هستند که ساختار سلولی ندارند، به همین دلیل برای تولیدمثل باید وارد سلول زنده شوند و با استفاده از مواد موجود در سلول میزبان تولیدمثل کنند.
ویروس در بیرون از ب*دن هیچ گونه فعالیت زیستی ندارد.
محتویات
- ۱ اجزای ویروسها
- ۲ ویژگیهای ویروسها
- ۳ آلودگی ویروسی
- ۴ دفاع ب*دن در برابر ویروسها
- ۵ موارد استفاده ویروسها
- ۶ منابع
- پوشش لیپیدی
- پوشش پروتئینی
- پروتئینها
- ماده وراثتی
- ویروسها ساختار سلولی ندارند. پیکر هر ویروس از یک بخش اسید نوکلئیک و یک پوشش درست شدهاست. تنها اسید نوکلئیک ویروس وارد سلول میشود.
- همه آنها برای تولید مثل حتماً باید وارد سلول شوند
- میزبان اختصاصی دارند، یعنی هر نوع ویروس فقط به یک نوع سلول زنده حمله میکند
- ویروسها به اندازهای کوچک اند که با الکتوسکوپ نوری نیز دیده نمیشوند و باید آنها را با میکروسکوپ الکترونی دید.
ویروسها تقریباً تمام موجودات زنده را مورد حمله قرار میدهند.
دفاع ب*دن در برابر ویروسها
هنگام حمله ویروسها به ب*دن، سلولهای مورد حمله مادهای پروتئینی به نام اینترفرون تولید میکنند. پس از پارهشدن سلولها و آزادشدن ویروسها، اینترفرون به سلولهای سالم میچسبد و آن سلولها را نسبت به ویروس بطور موقت مقاوم میکند.
موارد استفاده ویروسها
- کنترل تعداد حشرات مضر از راه بیمار کردن آنها توسط ویروسها
- تحقیق درباره DNA. کارهای آن در سلول
- کنترل بیماریهای ویروسی از راه تهیه واکسن از برخی ویروسها
موضوعات مشابه
- پاسخها
- 5
- بازدیدها
- 110
- پاسخها
- 7
- بازدیدها
- 118
آمار آنلاین
- کاربران آنلاین
- 11
- مهمانهای آنلاین
- 289
- مجموع بازدید کنندگان
- 300
مجموع ، شامل کاربران مخفی نیز میباشد.